|
|||||||||
|
|||||||||
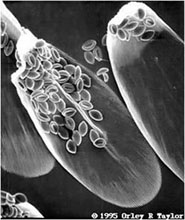
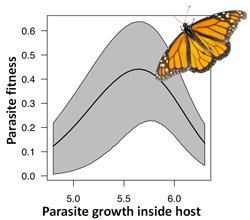
Evolution of parasite virulence: virulence-transmission trade-offsWhy do parasites - which rely on their hosts for survival - cause disease or even their hosts? A major evolutionary theory poses that parasite virulence can evolve as an unavoidable consequence of natural selection on parasite transmission. The idea is that parasites need to transmit between hosts and that higher host exploitation results in greater transmission. However, parasite growth also uses up host resources and damages tissues, and thereby results in disease and an increased probability that the host will die. Thus, high parasite growth inside a host has costs and benefits: the costs are increased disease and mortality, while the benefits are increased transmission to new hosts. On the basis of this trade-off it is expected that parasite strains with intermediate growth rates are selected for in nature: this is because these parasites grow fast enough to obtain transmission to new hosts, but not so fast as to kill the host before such transmission can occur. We have experimentally tested this theory, using the protozoan parasite Ophryocystis elektroscirrha, which infects monarch butterflies (Danaus plexippus) in natural populations. This parasite infects monarch butterflies at the larval stage, when caterpillars ingest parasite spores that are deposited by adult butterflies on the milkweed plants that these caterpillars eat. Parasites replicate to high numbers in the monarch caterpillar, such that adult butterflies can carry millions of parasite spores. In our work, we found that these high numbers of parasite spores are necessary to ensure parasite transmission to the next generation: transmission of spores occurs from egg-laying adult butterflies to their offspring caterpillars. |
||||||||
|
||||||||
|
||||||||
Effects of environmental factors on parasite virulence evolution |
||||||||
|
||||||||
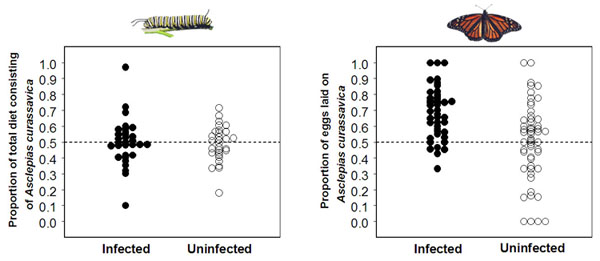
Ecology and evolution of self-medicationBecause parasites cause disease and death in their hosts, there should be strong selection for hosts to evolve ways to protect themselves against parasites. One of the most intriguing ways in which animals can protect themselves against disease is medication, through which animals use naturally available drugs to fight their parasites. Such medication has often been suggested to occur in great apes and other primates, but there remains a lack of strong evidence for medication in nature. Our lab recently tested for the existence of medication in monarch butterflies. Based on our findings that certain milkweed species can reduce parasite infection and growth and thereby relieve disease symptoms, we predicted that monarch butterflies would be able to specifically use such medicinal milkweed plants when infected. In a series of experiments, we found that monarch caterpillars cannot actively choose medicinal plants when infected. However, we found that infected female butterflies preferentially lay their eggs on milkweed that will make their offspring less sick. These results suggest that monarch butterflies have evolved the ability to medicate their offspring, and provide strong evidence that wild animals can use medication to fight their parasitic infections. Together with Mark Hunter’s lab at the University of Michigan we are working to identify the chemicals that make milkweeds medicinal. Because many parasites afflicting humans – including those that cause malaria, toxoplasmosis and cryptosporidiosis – are closely related to the parasites afflicting monarch butterflies, identifying these chemicals may provide new drugs against human parasites. As part of this work, we are currently testing the hypothesis that levels of disease risk determine the type of medication that animals evolve. In particular, we predict that when disease risk is high and predictable, animals will evolve a prophylactic form of medication ("take drugs before getting sick"), and that when disease risk is low, animals will evolve a therapeutic form of self-medication ("only take drugs when sick").
|
||||||||
Evolution of host defense against parasite infectionSelf-medication is just one of the many ways in which organisms may protect themselves against parasites. Other ways involve resistance (prevent infection or reduce parasite numbers) and tolerance (alleviate the symptoms of infection without preventing infection or reducing parasite numbers). We have studied both these mechanisms in monarch butterflies and found that monarchs vary genetically in resistance, but not tolerance. To better understand monarch defenses against parasite infection, we are currently working on a project to sequence the transcriptome of infected and uninfected butterflies. This will enable us to identify genes that are specifically expressed in monarchs infected with their parasites. We are doing this work in collaboration with Nicole Gerardo and Tim Read's Emory GRA Genome Center, both here at Emory.
|
||||||||
Ecology and evolution of infections of multiple parasite species
|
||||||||
Evolution of virulence in Varroa mites of honeybeesWe recently started to work with Berry Brosi (Emory) and Keith Delaplane (University of Georgia) to study whether current bee-keeping practices select for highly virulent bee mites. Honeybees are declining worldwide, and much of these declines are attributed to "Colony Collapse Disorder". However, Varroa mites pose a greater threat to honeybees, causing great natural and economic losses worldwide by wiping out large fractions of beekeeper' colonies. One explanation for the great harm done by these mites is that current bee-keeping practices are actually selecting for highly virulent mites by facilitating high levels of mite transmission between colonies, apiaries and different geographic regions. In theory, such facilitation of transmission should select for highly virulent parasites. We are currently testing this hypothesis by comparing the transmission and virulence of mites in managed and feral bee colonies. |
||||||||
Parasitoid avoidance in fruit fliesLike other organisms, fruit flies have many natural enemies, including parasitoid wasps that lay their eggs inside fruit fly larvae. Our colleague Todd Schlenke has studied these wasps for many years, and we have recently joined his research by studying whether fruit flies can behaviorally avoid wasp attacks. In particular, we are investigating whether fruit flies lay fewer eggs when surrounded by parasitoid wasps, and whether they can postpone their egg-laying until wasp threats have subsided. We are also investigating the mechanisms by which fruit flies detect the presence of wasps, using all sorts of cool mutants that will probably never be available for monarch butterflies.
|
||||||||
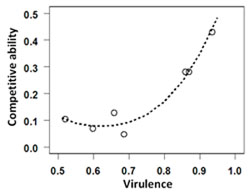
Effects of competition between drug-resistant and –sensitive malaria parasites on the spread of drug resistanceAs outlined above, parasite infections often consist of multiple parasite genotypes. In many cases, such mixed infections can contain parasites that are sensitive and those that are resistant to anti-parasitic drugs. Because parasites in mixed infections have to compete with each other for limited resources (see above), it can be expected that drug-resistant strains do not produce as many offspring and do not obtain as much transmission when competing with other strains than if they infected a host alone. Interestingly, if this is the case, then it can also be expected that the removal of drug-sensitive strains through the use of drugs removes this competition. This, in turn, could result in an increase in growth and transmission of the drug-resistant strain, and thereby aid in its spread through a population.
|
||||||||